
A Four-Letter Word Called “Race”
by Dr. C. Loring Brace
October/November 2000
Originally prepared for:
Race and Other Miscalculations, and Mismeasures:
Papers in Honor of Ashley Montagu
L. J. Reynolds and L. Lieberman (eds.)
General Hall, Publishers
Dix Hills, N.Y.
Prologue

The very genesis of the field of biological anthropology was rooted in the assumption that there are valid biological entities that can be called “races,” and that these then are legitimate targets of anthropological study. Biological anthropologists have often been somewhat slower than others to realize that those “entities” do not have coherent biological reality. This has been particularly true in America where the concept of “race” was more an ancillary result of the circumstances of the human settlement of the hemisphere and how this affected the way people think about things than a consequence of the study of the real nature and dimensions of human biological variation.
The first biological anthropologist in America to bring this to the attention of the reading public was a transplanted Englishman, Ashley Montagu, who settled in the United States in 1930, the very year that I was born, and has remained here ever since (Brace, 1997). Montagu’s most influential work, Man’s Most Dangerous Myth: The Fallacy of Race (Montagu, 1942), has had a turbulent history. In the past, much of biological anthropology tended to treat it as though it were a manifestation of what we now refer to as P.C. — political correctness — in this case a kind of ‘feel-good’ manifestation of liberal wishful thinking (for example, Shipman, 1994; but see the critique of that by Brace, 1995).
The approach may indeed be compatible with liberal social thought, but it is actually based on a solid grasp of the biological nature of human variation. As the synthetic theory of evolution began to have its impact on some parts of biological anthropology in the decade after the end of World War II, there were some who realized that we could only make real biological sense out of the nature of human variation after the concept of “race” was junked and we started over from scratch. I was one of the people who came to that realization when I started teaching at the Santa Barbara campus of the University of California. I had been able to get Ashley Montagu to speak to our undergraduate anthropology club there, and I was successful in getting him brought to the campus in 1963 as Regent’s Lecturer for our winter term. In turn he invited me to contribute a chapter for the volume, The Concept of Race, that he was editing to appear in 1964 (Brace, 1964). In that chapter, I tried to show how to handle the study of the nature of human biological variation after the “race” concept had been dispensed with.
That was a third of a century ago, however, and there has been more than a bit of back-sliding in the world of biological anthropology. With that in mind, Larry Reynolds and Leonard Lieberman, of Central Michigan University, prepared a volume of essays in honor of Ashley Montagu’s ninetieth year, and, to my great pleasure, they invited me to be one of the contributors. This gave me the opportunity to completely revamp and update the approach I had taken in 1964 with the addition of the quantities of information and insight that have accumulated since that time. Actually since so many biological anthropologists still have not gotten much beyond the point of trying to justify the application of “racial” names to human individuals and groups, the information relating to the distribution of the separate aspects of human adaptation has not accumulated as rapidly as it could have. Hope springs eternal, and, this time around, just maybe the idea will take root and a new generation will actually go and get the information we need to document the picture of which I present here in outline form (Brace, 1996).
When I presented a version of this to the Canadian Association of Physical Anthropologists in Windsor, Ontario, on October 28, 1994, I was able to get the assistance of Humbert O. Echo, another member of that extraordinary faculty in the Department of Homopathic Anthropopoetics at the University of Southern North Dakota at Hoople. Echo had previously been of assistance by rendering some of the gambits I have essayed in versified form, and he had allowed me to use that particular piece as an addendum to a comment, “What shall we call ‘Them’?” that I published in 1996 (Echo, 1996). This is attached here:
The Name of a Race
When we ponder on the contours in the features of a face
That resembles all the others which are from a given place;
What potential harm would follow if we use a single name,
To denote a group of people when we think they look the same?
But there are no implications that a common shape will bear,
Beyond the clear reminder of the kinship that they share;
For selection’s not delimited by groups of kin alone,
Or confined within the boundaries of a continental zone.
Pigment in the skin will give protection from the sun,
But it doesn’t give a clue to how another trait will run;
But the desert and the artic take the moisture from the air,
And people from both places have a nose of length to spare.
Features cannot tell us who is mad and who is sane,
Or nuance of the forehead say a thing about the brain.
Each trait that is adaptive will pursue a separate course,
Determined by the nature of its own selective force
Which crosses all the others in a fashion that defines
A pattern without meaning made of independent clines.
Since each selected feature has a different place of birth,
The mix within a region can have no collective worth.
When thoughtlessly we verbalize without the proper care,
Our words can make an entity that isn’t really there;
How much pigment or how little will suffice to give the right
To warrant the conferral of the label “Black” or “White”?
And beware the added meaning in the tag we lightly give;
For it oftentimes determines who may have the right to live.
Acceptance of the concept, and all that it can mean,
Gives credence to an image that could best be called obscene;
To use the very word is to be captured by its spell:
That which we call a “race,” by any other name would smell…
Sources Cited
Brace, C. Loring, 1964. A Non-Racial Approach Towards the Understanding of Human Diversity. In Ashley Montagu (ed.), The Concept of Race. The Free Press of Glencoe, New York. pp. 103-152.
Brace, C. Loring, 1995. Review of The Evolution of Racism: Human Differences and the Use and Abuse of Science, by Pat Shipman. American Journal of Physical Anthropology 96(2):204-210.
Brace, C. Loring, 1996. A Four-Letter Word Called Race. In Larry T. Reynolds and Leonard Lieberman (eds.), Race and Other Misadventures: Essays in Honor of Ashley Montagu in His Ninetieth Year. General Hall, Publishers, Dix Hills, New York. pp. 106-141.
Brace. C. Loring, 1997. Montagu, Ashley (1905- ). In Frank Spencer (ed.), History of Physical Anthropology: An Encyclopedia. Garland, New York. pp. 683-685.
Montagu, Ashley, 1942. Man’s Most Dangerous Myth: The Fallacy of Race. Columbia University Press, New York. 216 pp.
Shipman, Pat, 1994. The Evolution of Racism: Human Differences and the Use and Abuse of Science. Simon & Schuster, New York. 318 pp.
Abstract
Human biological variation is real, and its study is a most interesting subject. However, we can make no sense of it if we start with “racial” categories as entities for comparison. Traits that are of importance for human survival are distributed in clinal fashion according to the distribution of the selective forces that govern their expression. Those selective forces, in turn, are not constrained by human population boundaries and cannot be perceived or understood if such boundaries are assumed as the starting points for analysis.
There are traits that are constrained by human population boundaries, and it is these that allow us to recognize the part of the world from which their possessors originally came. Those traits, however, simply constitute ‘family resemblance writ large’ end have no adaptive significance. Population samples will cluster with their nearest neighbors because they share the genetic background for such adaptively unimportant traits. The best way to refer to people who display the configuration characteristic of the clusters of population samples that come from the same region is to use geographical designations. Thus people can be identified as African, or Australian, or European and the like. More precise localization can be achieved by using modifiers such as West African, Eastern European, and Southeast Asian among any others.
Data relating to the distribution of Hemoglobin S, skin color and tooth size are presented to show how each responds to the varying intensity of the particular selective force controlling its manifestation. The independence of the clines associated with each one demonstrates the futility of trying to use a concept such as “race” to understand adaptively important human biological characteristics. Finally, the case of intelligence is considered, and it is noted that, while no human population today is living under the circumstances that shaped the common human condition during the Pleistocene, it is still largely true that it takes at least as much intelligence to survive and contribute to the next generation in one part of the world as it does in another. Since the conditions governing the emergence of our extraordinary human brain power were essentially everywhere the same during the long period of time when human intelligence was evolving, it follows that the intellectual capabilities of the various living human populations in the world are now also everywhere the same. Those who continue to advocate the value of investigating “racial” difference in intelligence, then, are evidently driven by a subjective desire to demonstrate that difference is to be expected. At bottom, that expectation is simply a manifestation of racism.
Introduction
All good people agree,
And all good people say,
All nice people, like us, are We
And everyone else is they.
Rudyard Kipling, We and They
The issues dealt with here are so important that they should transcend the involvement and pronouncements of any single individual or school of thought. If there is a possible exception to that generalization, it would have to be illustrated by the life and work of Ashley Montagu For more than half a century now, he has stood for the view that “race” is not only a “myth,” but that it is our “Most Dangerous Myth,” to quote from the title of one of his most influential books first written in 1942. The insights in that seminal work remain as true today as when he first phrased them, and they can serve as a solid point of departure for my own efforts in this paper written to celebrate the career and accomplishments of the author of that signal contribution. This gives me the opportunity to update my own first attempt (Brace 1964) to deal with the question of “race” written some thirty years ago at the specific request of the very person in whose honor I am writing now — Ashley Montagu.
I am going to begin with the conclusion: “race” is a social construct derived mainly from perceptions conditioned by the events of recorded history, and it has no basic biological reality. Quite simply, there is no useful entity that corresponds to what is popularly intended by the term “race.” This was explicitly noted in regard to the case of Americans of African ancestry by no less a figure than the late Gunnar Myrdal — that extraordinary Swedish economist and author of one of the most perceptive books ever written about the way things are in the United States, An American Dilemma (1944:115). To much of the reading public, this will seem like a complete absurdity. The average literate citizen of the western world reacts with frank disbelief when told that there is no such thing as “race.” “Why, it’s as plain as the nose on your face!” is one of the partially facetious reactions. When the anthropologist continues to insist that there is no human biological category that can be called “race,” the skeptical layman will shake his head and just regard this as further evidence of the innate silliness of those who call themselves intellectuals.
This feeling has been seconded by some biological anthropologists who have gone so far as to say that denials of the biological reality of “race” are simply the products of their well-meaning colleagues’ abhorrence of the ills and injustices that have arisen in its name. In this view, the denial that “race” has a biological reality is itself the result of socially conditioned perceptions. Myrdal, for instance, was a social scientist and not recognized as an authority on biological matters. However well-intentioned they may be, then, such denials themselves presumably have no biological justification and therefore lack validity. The cry of both professional and non-professional skeptics goes like this: ‘If there are no “races,” how come people are so good at identifying them?’ (and see how that rhetorical question is asked — and answered! — in Sauer 1992).
But what is this “them” that we are so good at recognizing? It is true that people are reasonably good at being able to tell what part of the world someone comes from in a general kind of way. Unless a person has parents from very different parts of the world, it is not hard to tell whether a person’s family roots were in eastern Asia, western Europe, or southern India. In addition, most Americans would feel confident that they could detect the presence of African ancestry, although in fact they are quite unable to tell whether a person comes from West Africa or the eastern end of New Guinea. There are reasons for that particular confusion which will be treated later, but, although it introduces a touch of uncertainty, it does not really blunt the general conviction that it is not all that difficult to tell at a glance the part of the world from which a person’s ancestors originally came.
Traditional “Races”
So far, however, I have said nothing about “race” — I have just mentioned the characteristic appearance associated with geographic areas. Well, you may ask, why isn’t that recognizable appearance an indicator of the presence of something legitimately called “race”? To answer that, we have to come to grips with what it is that produces those configurations that we can associate with given areas, and we have to consider the biological significance of each such configuration.
One of the most enduring schemes of “racial” designation divides the peoples of the world into three large categories crudely conceptualized as “black,” “white,” and “yellow;” or in more orotund and polysyllabic form, “Negroid,” “Caucasoid,” and “Mongoloid.” Early in the 19th century, this scheme was advocated by Georges Cuvier (1769-1832), one of the most influential figures in the history of French science, although it was neither original with him nor was it offered in anything more than the most casual of terms (Cuvier 1817:1:94; 1829:1:80). Some of Cuvier’s readers interpreted these three “races” as the respective descendants of the three sons of the Biblical Noah — Ham, Shem, and Japheth (Murray 1834:255; Morton 1839:2).
For many of the beneficiaries of the traditions of “western civilization,” this formulation was eminently satisfying because it drew strength from its apparent roots in the Bible, the most honored written work in existence in the minds of the Christian faithful who made up the overwhelming majority of the representatives of that “western civilization.” In addition, it evoked the sanctity of what westerners regarded as that most mystically sacred of quantities, the number three — also long associated with the Christian “Trinity” even though that baffling concept can only be vaguely and somewhat tortuously squeezed out of the actual phraseology of the Bible.
Now let us take a closer look at what it is that makes us think that we can understand something about human variation if we divide the world up into those three categories. Are they really comparable in the sense of being equally distinctive? And is there something about each that justifies its identification as a category, and what is the biological significance of that “something” if indeed it exists?
Negroid
The very name “Negroid” derives from the Latin word for black — niger — and the word used to denote that color in modern Italian, Spanish and Portuguese is negro. Because southern Europe is closer to Africa than the other European countries, it was inevitable that contacts between the peoples of southern Europe and Africa began earlier and were carried on more continuously than was true between Africans and more northerly European nations. Eventually when northern Europeans extended their contacts towards the south, they tended to adopt the terms already in use by their Mediterranean neighbors. Early in the history of English involvement in the growing trade of enslaved Africans, and long before there were any English-speaking settlers in the western Hemisphere, the term “Negro” entered the English language to mean a native inhabitant of the African continent south of the Sahara (Pope-Hennessy 1968:46). The identification of native Africans as “Negroes” served to focus the attention of would-be categorizers on a single descriptive attribute — skin color. The presence of sufficient melanin in the skin to warrant using a term such as “Negro” or “Black,” however, is not restricted to the people of Africa alone. The native inhabitants of southern India, New Guinea and adjacent islands, and northern Australia all possess equivalent amounts of melanin in their skins. If a word meaning “Black” is warranted as a description for people of African origin, then it is also equally appropriate for those others.
The use of the term “Negroid” obviously cannot distinguish between the long-term equatorial inhabitants from one end of the Old World to the other. In fact, melanin in the skin is an adaptation that shields the possessor from the damaging effects of the ultraviolet component of sunlight, and all human populations whose immediate ancestors had been continuous dwellers of the tropics for 100,000 years or more possess that adaptation to an equal extent even if they are only remotely related to each other. As Darwin explicitly realized, any trait that is under the control of selective forces is “almost valueless” for purposes of tracing population relationships (Darwin 1859:427). Obviously a classification based on a trait whose manifestation is under the control of selection will tell us much about the distribution of the relevant selective force, but it will tell us little about the degree of relationship of those populations that display similar degrees of development of the trait in question.
Traditionalists unwilling to give up so soon may raise points concerning the presence of other traits visible in people with elevated levels of melanin in the skin, and suggest that these hang together to indicate the presence of some kind of fundamental underlying entity. Dark skin tends to be associated with tightly curled hair, for example, and dark-skinned people often have large jaws and teeth. However, hair form is the product of the same kinds of selective forces that are associated with skin color. Jaws and teeth, on the other hand, vary according to quite different rules as we shall see later. For the dark-skinned people of New Guinea and Australia, for example, jaw and tooth size actually increases as the intensity of skin pigmentation decreases grading from the North of Australia down to its southern edge (Brace 1980; Brace et al. 1991). Clearly the idea of a “Negroid race” has so many flaws that it is best simply to drop it and start on another tack. More of that later.
Caucasoid
All right, you may say with some reluctance, if an essentially descriptive word does not work to categorize human populations, how about a word that is relatively abstract — “Caucasoid” for example? What could be wrong with using that presumably innocuous term to stand for the people in the northwestern quadrant of human habitation? The history of the application and use of the term “Caucasoid,” however, introduces still further problems. The word derives from the Caucasus, that isthmus of land that separates the Black and the Caspian Seas and joins Russia and the Middle East. Its use to denote human physical appearance dates from the “racial” scheme offered by the German physician, Johann Friedrich Blumenbach (1752-1840), in his doctoral dissertation of 1775, De Generis Humani Varietate Nativa (On the Natural Varieties of Mankind) best known from the revised and enlarged third edition of 1795 (translated and edited by Thomas Bendyshe in 1865 and republished in 1969).
The initial reason for the focus on that part of the world was the assumption that, as the Biblical flood subsided, Noah’s Ark landed at Mount Ararat in the mountains of the Caucasus. Although the idea was not original with him, Blumenbach claimed that the living people of that region comprise “the most beautiful race of men” with “the most beautiful form of the skull, from which, as from a mean and primeval type, the others diverge by most easy gradations on both sides” (Blumenbach in Bendyshe [ed.] 1865:269). Adding to this assumption of pristine beauty, Blumenbach went on to claim that “white . . . we may fairly assume to have been the primitive colour of mankind” (idem.).
Blumenbach evidently assumed that “Caucasoids” were the least modified descendants of the people who allegedly got off the Ark at Mount Ararat and therefore the best representatives of what God intended when He created human beings in the first place. Other “races” were said to have departed from the form of God’s original intent by a process of “degeneration” in proportion to their geographical distance from the mountains of the Caucasus and the differences in the circumstances under which they now find themselves.
I have actually written an overly simplistic synopsis of Blumenbach’s thoughtful treatise, and I have left out many of the positive things that it contained. My purpose is not to fault Blumenbach but to highlight the absurdity of the term “Caucasoid.” One could go even further by noting that Mount Ararat is actually in Turkish Armenia, and, if there were any justification in using point geographic designations to refer to broadly related human populations, it could be argued that the term “Armenoid” should be preferable to the term “Caucasoid.” Of course, the designation “Armenoid” was used for somewhat different purposes in other “racial” classifications a couple of generations ago (Coon 1939:628-629), and this makes it as tainted as “Caucasoid.” Again, the best thing to do is to abandon the use of any narrow regional designation as a means of encompassing the perceived similarities of human groups distributed across widespread geographic expanses. The related morphological pattern that we can see running from Scandinavia to the Middle East is poorly served by trying to indicate this by using a narrowly local term, whether that be “Armenoid,” “Norwegioid,” or “Caucasoid.”
Mongoloid
All right, so descriptive terms and narrow regional designations fail to serve our purposes, but what about names derived from human groups? What could be wrong with the use of the term “Mongoloids” to refer to all the related people of eastern Asia? In fact, it has all of the flaws present in narrow regional designations plus some others as well. Just as the Norse or Armenians are inappropriate to use as representatives of all of the people who extend between Norway and Armenia, the Mongols are inappropriate to stand for all of the people who extend from Mongolia to Indonesia.
Adding further less-than-positive connotations to the term “Mongoloid” is the long-time usage of that term to refer to the visible features associated with an inherited syndrome that occurs as a result of irregularities in the transmission of chromosome 21. The first full description was published in 1866 by the English physician, John Langdon Down (Patterson 1987).
When the syndrome recognized by Dr. Down was first described, it was noted that the English children in whom it was observed were afflicted with developmental defects that influenced a series of systems. These included the growth and ossification of the interorbital portion of the face as well as the normally expected path of intellectual maturation. Affected children are also observed to display an alteration in the expected course of pigment production giving a slightly yellowish cast to their appearance. In the European mind, all of these manifestations reminded them of their stereotypic picture of the inhabitants of eastern Asia — flattish of face, yellowish in color and mentally dull and lethargic. The inherited interference with development observed by Dr. Down then was duly labeled “Mongolism” or, in more pejorative fashion, “Mongoloid idiocy.” Implicit in this was the feeling that the genetic defect which caused the syndrome was related to the hereditary reasons why the people of eastern Asia differ in general appearance from the people of western Europe. Subsequent generations of geneticists and physicians have recognized the blatant racism behind such attitudes and have sought to correct this by eliminating the term “Mongoloid” from the description of that suite of inherited developmental defects and describing it simply as “Down’s syndrome.” More recently, the possessive has been eliminated, and it has come to be called “Down syndrome” or “Down disease,” and no mention is made of the supposed similarity in appearance between those who are afflicted and the inhabitants of Asia. The memory lingers on in a large segment of the public in the western world, and, if for no other reason, this alone would make the use of the term “Mongoloid” an unfortunate choice of words to describe the population of a major segment of the earth.
However, there is more than just the taint of that racism in the terms of yesteryear to illustrate the fact that the designation “Mongoloid” is misleading at best when used in descriptive fashion. The Mongols, as it happens, display systematic morphological differences from the rest of the people in Asia to such a degree that they could be regarded as the most untypical of that whole set of related populations (Li et al. 1991:278). This is not just an offhanded judgment. A standard set of two dozen craniofacial measurements (the measurements and the techniques used are described at much greater length in Chapters 19 and 20) was made on samples chosen from all the available localities in eastern Asia. These were used to construct a tree diagram — the “dendrogram” shown in Figure 1 — where the length of the stem of each twig is proportional to the morphological relationship with the other twigs to which it is connected. A short stem running from a named twig to the next connecting link means a close relationship. The longer the stem on the twig, the more remote the relationship.
In Figure I, the twig connecting the Mongols to the rest of the samples from eastern Asia is the longest unlinked branch in the entire dendrogram. Other studies using slightly different techniques have come to the same conclusions (Pietrusewsky 1992:42; Pietrusewsky et al. 1992:552). Mongols, then, are as morphologically remote from the other inhabitants of eastern Asia as it is possible to be and yet still count as related (Brace and Tracer 1992:459). To use the Mongols as though they could stand for something essential to all of the related populations of eastern Asia is to run the risk of serious misrepresentation at best. Of course, the assumption that there is some sort of “essence” that we should be looking for is itself fundamentally misguided.
Clines
A generation ago, Frank Livingstone produced the aphorism, “There are no races, there are only clines” (Livingstone 1962:279). This fit in nicely with the assumptions of the neo-Darwinian or synthetic theory of evolution that characterized much of the outlook of the biological sciences that arose in the middle third of the twentieth century. One of the most important figures in that synthesis was the late Sir Ronald A. Fisher (1890-1962), an English biological statistician whose quirky genius occasionally led him into positions of dogmatic intransigence (Box 1978). His long-running feud with the American geneticist, Sewall Wright (1889-1998), was a case in point.
Wright had entertained the possibility that biological evolution could be conditioned by a number of different factors including chance alone — especially when population size is small (Wright 1931). Fisher simply refused to acknowledge the actual mechanisms Wright proposed (Fisher and Ford 1950; Wright 1951). Their argument narrowed down to whether genetic drift could or could not play a role in addition to the effects of natural selection. Fisher had taken the view that selection is all (Fisher 1930), and that in those instances where we could not see how it had its effect, the fault was in our finite and limited powers of observation and understanding. Biologists who accepted this conclusion were put in the position of assuming that all discernible inherited traits and configurations owed their various manifestations to the controlling effects of selection. Our duty, then, was to determine the nature of the selective forces involved and to demonstrate just how these work to control variation in the particular traits that respond to changes in the intensity of their operation. If all traits respond to the graded influence of their particular selective forces, the intersection of their various manifestations to make identifiable configurations in given individuals or populations has no intrinsic significance. This in brief is the intellectual background behind the phrase “there are no races, there are only clines.” Whenever separate adaptively significant traits are controlled by separate and unrelated selective forces, this still remains true.
Hemoglobin S
Let me give a concrete example using variation in human populations. Before Livingstone coined that portentous phrase, he had shown that the frequency of the gene for hemoglobin S — the source of human sickle-cell anemia — is controlled by the intensity of infestation with falciparum malaria (Livingstone 1958). Populations in the malarial areas of tropical Africa, the Middle East, and on into India all show high percentages of hemoglobin S even though a certain number of S genes are taken out of circulation each generation because of deaths due to sickle cell anemia.
The distribution of hemoglobin S, then, is unrelated to human population boundaries. It is not an automatic marker for African ancestry as has often been assumed, although many of the Americans who possess it did inherit it from African ancestors. Africa itself probably received hemoglobin S from the Middle East via Arab trade routes across the Sahara, up the Nile, and down the east coast (Livingstone 1989b). The circumstances that led to the infestation by Plasmodium falciparum, the mosquito-transmitted microbe that causes falciparum malaria, are associated with the increase in human population size and the environmental modifications produced by successful agricultural practices in areas where that malaria-producing parasite can flourish. Those areas originally included the tropical, subtropical and temperate parts of the Old World with enough water to allow propagation of the mosquito vector and where the winter was not so severe that they could not survive from one season to the next.
Figure 2 shows the distribution of hemoglobin S in the Old World. In its areas of highest frequency, falciparum malaria also reaches high levels of intensity. If falciparum malaria is the reason for the high frequency of hemoglobin S, one might expect that there would be a one-to-one correlation between malaria intensity and hemoglobin S, but this is not exactly the case. In Southeast Asia, for example, falciparum malaria constitutes a very serious health problem, but hemoglobin S fades out and disappears. The reason for this is the presence of another abnormal hemoglobin — hemoglobin E — which, as it happens, also enables its possessors to cope with malarial infestation but which does not have quite such serious consequences in its homozygous — EE — state (Livingstone 18998a).
Skin color
The abnormal hemoglobins, then, are distributed among the world’s populations strictly in accordance with the history of their involvement with malaria of various kinds and without reference to any other manifestations such as geographical, cultural, or political boundaries or anything like face form or pigmentation (Livingstone 1967, 1985). In fact, this is the classic pattern for the distribution of any trait under selective force control. And when we turn to a consideration of pigment, that too shows a pattern of gradation which is independent of the distribution of any other trait not directly related to the intensity of solar radiation. Figure 3 shows the distribution of skin color in the human populations of the world.
Melanin in the skin exists for one purpose only, and that is to prevent the penetration of the ultraviolet component in sunlight. Attempts to suggest that melanin has any relation to mental function — pro or con — are nothing less than unwarranted manifestations of racism (Ortiz de Montellano 1991, 1992). However, it is the 290 to 320 millimicron ultraviolet range, “mid-UV,” “middlewave UV” or “UV-B” —.either alone or, as is usually the case, in conjunction with UV-A — that causes the most trouble (Potter 1985; Kligman 1986). The “trouble” caused by UV-B is cancer. Particularly affected are the cells at the bottom of the epidermis at the interface between this layer and the underlying dermis itself. These cells give rise to the outermost portion of the skin in a continued process of renewal that has a three- to-four week turnover (Daniels et al. 1968). UV-B can cause damage that leads to cancer in the basal cells and the derived overlying squamous cells that form the surface of the skin.
In the United States alone in the late 1970’s, European Americans contracted over 400,000 cases of basal cell skin cancer which amounted to an incidence of over 232 cases per every 100,000 people. At the same time, less than three-and-one-half cases per 100,000 occurred among Americans of African origin (Ackerman and del Regato 1985:180). At that time, the average number of Americans who died per year from the effects of skin cancer was some 5,000 (interview with Dr. Tim Johnson on ABC television, April 10, 1978).
Carefully designed and controlled laboratory tests have clearly demonstrated the role played by UV-B in leading to skin cancer and have shown that the effects are augmented when both UV-A and UV-B are involved, which of course they are in the case of people and animals exposed to natural sunlight. It has also long been known that “white” skin will allow the penetration of ultraviolet radiation right down through the epidermis to the underlying dermis (Daniels 1969; Kligman 1969; Pathak and Stratton 1969), but that the skin of well-tanned or naturally pigmented individuals will block up to 95% of the penetration of UV-B (Daniels et al. 1968:41; Holick 1987:1879). And in the occasional person of African ancestry in whom a mutation has occurred that blocks the formation of skin pigment — a condition known as albinism — the skin will allow the penetration of ultraviolet radiation in the same manner as in “whites.” African albinos are susceptible to skin cancer in the same manner as Europeans (Blum 1959; Friedman et al. 1991:10). From the accumulation of the experimental and clinical evidence, it is impossible to avoid the conclusion that the sole function of melanin in the skin is as the first line of defense against the possible cancer-producing effects of ultraviolet radiation from the sun.
That being the case, the distribution of human skin pigmentation in the world should correspond to the distribution of the intensity of ultraviolet radiation, or, roughly speaking, to latitude: i.e. dark skin color should be concentrated in the tropics where it is adaptively advantageous. From Figure 3, it is clear that the dark- skinned populations of the world are all located in the tropics. However, it is also clear that not all tropic dwellers are dark skinned. The people of mainland Southeast Asia and most of island Indonesia plus the remote islands of Oceania are nowhere near so heavily pigmented as the indigenous populations of tropical Africa, southern India, New Guinea or northern Australia. And finally, there were no indigenous dark-skinned people in the tropics of the western hemisphere.
The obvious reason for this apparent discrepancy is that the relatively light-skinned inhabitants now in those tropical regions must be comparatively recent arrivals. This suspicion is amply confirmed by the evidence available from history, linguistics and archaeology (Benedict 1942; Barth 1952; Bellwood 1979, 1986, 1991; Solheim 1981; Bulbeck 1982). Historical records, for example, show that there has been a continuing movement of peoples southward from the older centers of settled agriculture in China to what had been uncut forest lands in Southeast Asia. Even where there are no written records, the oral traditions of a northern origin and the trail of related languages all tend to confirm this general picture. The web of what is obviously recent linguistic divergence extends all the way out into the islands of the Pacific where archaeology tells us that settlers had reached the Philippines 5,000 year ago but had not gotten as far as Easter Island and Hawaii in Remote Oceania until about 1,500 years ago (Bellwood 1986:107 ff., 1991; Pawley and Green 1973; Tuggle 1979:189).
As for the western hemisphere, although the crucial events occurred long before writing was thought of, and so far back that linguistic evidence is equivocal at best (Nichols 1990:513; Greenberg and Ruhlen 1992:94), archaeology tells us that the first inhabitants came over from the northeast edge of Asia by the end of the Pleistocene almost 12,000 years ago (Haynes 1982:383; Hoffecker et al. 1993:51). The possibility still exists that people may have reached the New World somewhat earlier, but the signs are all tenuous and equivocal. From 11,500 years ago on, however, the evidence is clear and widespread, and it leaves most professional students of the matter with the reasonably comfortable feeling that the ancestors of the “Native Americans” did not arrive in the western hemisphere much before 12,000.
The ancestors of the modern inhabitants of the tropics in Southeast Asia, Polynesia and South America all came from the non-tropical portions of eastern Asia no earlier than the latter part of the Pleistocene. We can assume, then, that the skin color of those ancestors was not significantly different from that of the inhabitants of the temperate parts of eastern Asia today. The reason why they should have been characterized by that particular hue is a different matter that will be considered shortly. For the moment, however, what this tells us is that it takes a lot longer than 10,000 years for the selective effects of UV-B in tropical sunshine to produce the intensity of pigmentation seen in the skin of such modern people as the inhabitants of New Guinea or tropical Africa.
The related question is why people who live in the northern portions of the world have as little skin pigment as they do. All the available fossil and archaeological evidence indicates that ultimate human origins were in the tropics of Africa, and that even after spreading out from that base at the end of the lower Pleistocene a million years ago, humans remained tropic dwellers with only temporary northward forays for much of the rest of their existence (Brace 1991b, and in press). Humans still have the basic physiological characteristics and responses of tropical mammals, and we can guess that the common tropic-dwelling human ancestor of half a million years ago had a degree of skin pigmentation fully comparable to that found today among the inhabitants of equatorial Africa, India and New Guinea. This brings us back to the question of why some modern people are less heavily pigmented than others.
Since all of the long-term inhabitants of the north temperate zone are markedly less pigmented than long-term tropic dwellers, and since we can assume that the common human ancestor was dark, it follows that there must be something about living north of the tropics that leads to a reduction in skin pigment. There are in fact two possible explanations. One suggests that a lessening in the amount of pigment is adaptively advantageous in the north, and the other argues that depigmentation is simply the result of a reduction in the intensity of selection for the maintenance of epidermal melanin.
Those who prefer to see pigment reduction as the result of the positive action of selection point to the evidence indicating that heavily pigmented people are more likely to suffer the effects of vitamin D deficiency in the north temperate zone than is true for people with lesser amounts of skin pigment (Holick 1987). As it happens, those very UV-B wave lengths that lead to skin cancer after prolonged doses are also an essential part of the process that leads to the synthesis of vitamin D. The absorption and incorporation of calcium necessary for proper bone growth is mediated by vitamin D, and a lack of that crucial substance will result in abnormalities and stunting of skeletal development known as rickets.
Rickets, however, is a distinct rarity among the cats, dogs, cows, horses, sheep, rodents, birds and others of the animal world that were native inhabitants of the north temperate zone in spite of the fact that few if any indulged themselves with codfish livers and that the skin in all of these creatures is well protected from any possible ultraviolet penetration by virtue of being thoroughly covered by fur or feathers. There is one more oddity about this whole picture, and this is the fact that there is no pigment in the skin of those thoroughly protected creatures. And finally, it only requires a brief exposure to generate all the vitamin D one needs, and summer sunshine in the north temperate zone is more than adequate even for heavily pigmented skin to provide a supply of Vitamin D that can be stored up in fat and muscle tissue in sufficient quantities to last the rest of the year (Robins 1991:203,205,208).
From my perspective, it would seem that the reason for the absence of pigment in the skin of most furred and feathered creatures is because of the absence of selection for its presence. And I would argue that the same thing may well be true for the human inhabitants of the north temperate zone. Since there is just not enough intensity of UV-B to generate cancers that would shorten possible life spans, there was nothing that would maintain skin pigment at the level which had characterized the tropical forebears of the long-term human inhabitants of the north. A generation ago, I argued that when selection is reduced or suspended, any trait that had been maintained by a formerly active selective force should undergo reduction in proportion to the length of time that selection had been held in abeyance. Under such conditions, the reductions would be the results of mutations alone — a process I referred to as “The Probable Mutation Effect” (Brace 1963).
A more detailed defense of the “PME” is presented elsewhere (Brace et al. 1991 :41-46, and included here as Chapter 10). Here we need only note that those areas where the archaeological evidence indicates that human habitation of the north temperate zone has been continuous for the longest periods of time are also just those areas where modern human populations display the least amount of pigment in the skin. In a zone running from the Middle East to the Atlantic Ocean in northwest Europe, there is reason to believe that human habitation has been continuous for the last 250,000 years (Straus 1989; Mercier et al. 1992). It is in just that stretch where the living inhabitants include people with the least amount of pigmentation among the living populations of the world.
At comparable latitudes in the northeast end of human occupation in the Old World, the continuous archaeological record is only about half as old. From this, it would follow that relaxation of selective pressures to maintain skin pigmentation at fully tropical levels has been in effect only half as long, and depigmentation should not have proceeded to the same extent as in the comparable latitudes in the west. Indeed, the modern inhabitants of Northeast Asia are notably less pigmented than the populations who have continued to live in the tropics without break, but pigment reduction has not proceeded to quite the same extent as that visible at the northwestern edge of the human range. And finally, it only requires a brief exposure to generate all the vitamin D one needs, and summer sunshine in the north temperate zone is more than adequate even for a heavily pigmented skin to provide a supply of Vitamin D that can be stored up in fat and muscle tissue in sufficient quantities to last the rest of the year (Robins 1991:203, 205, 208).
Actually, it does not matter which attempt at explanation is correct since both propose reasons why pigment reduction should occur in areas where ultraviolet radiation is markedly reduced from the levels of its intensity in the tropics. Either would account for the existence and extent of depigmentation in the north temperate zones both east and west in the Old World, and both offer reasons why the relatively recent movement of people from the latitude of China south into Southeast Asia and equatorial Indonesia introduced people with lesser amounts of skin pigmentation than one would otherwise expect to find for people living in the tropics.
At the same time, the movement of agriculturally based populations out of the Middle East during the Neolithic and Bronze Age was predominantly an East-to-West phenomenon rather than a North-to-South one (Brace and Tracer 1992). The result was that the North-to-South gradation or cline in skin color was not disrupted. To this day, skin color grades by imperceptible means from Europe southwards around the eastern end of the Mediterranean and up the Nile into Africa. From one end of this range to the other, there is no hint of a skin color boundary, and yet the spectrum runs from the lightest in the world at the northern edge to as dark as it is possible for humans to be at the equator.
South of the equator, the cline reverses, and skin color becomes lighter away from the tropics towards the south. In Africa, this lightening is more pronounced than the comparable case in Australia, and the aboriginal inhabitants of the southern tip of Africa — the San people once called by the derogatory term “Bushmen” — are no darker than the people by the shores of the Mediterranean who are about the same distance north of the equator as the San are south of it. Skin pigment also lightens towards the south in Australia, but not quite to the same extent as in the African example. Australia, however, has only been occupied for the last 50,000 years (Roberts et al. 1990), and evidently the process of pigment reduction has not had time enough to proceed to the extent evident at the comparable latitudes in Africa.
From all of this, it can be seen that human skin pigmentation is distributed in clinal fashion among those people who have remained in the latitudes where they are found for a period of time on the order of 50,000 years or more. Where skin pigmentation is at variance with our expectations of clinal variation such as Southeast Asia and the New World, we have reason to suspect that such anomalies in human appearance are due to population movements within the last 20,000 years or so. So far, all such suspected instances are confirmed by the available archaeological evidence.
Finally, it is clear that the distributions shown in Figures 2 and 3 are following different sets of rules. Evidently neither one can be understood if one uses the old- fashioned concept of “race” as a starting point, and yet both display vital but unrelated aspects of human biological variation. Each evidently is clinal in nature, but the clines have little to do with each other. Each can be readily understood when studied separately, but the pattern made by the intersection between these two has no “meaning” in and of itself. And when one adds a third trait or a fourth or more, the configurations that emerge tell us nothing whatsoever about the nature and significance of human biological variation.
Tooth Size
The third trait that I am going to deal with is tooth size. It is neither more instructive nor in other ways “better” than any other trait one could use, but it serves the purpose, and I can use the information that I have been collecting myself for the past several decades to demonstrate the nature of modern human tooth size differences.
There has long been a vague kind of perception on the part of European observers that “other races” had larger jaws and teeth than those found in Europe. Along with this perception of greater size in jaw and tooth was a related assumption of smaller size of head and brain. These were interpreted in a hierarchical fashion which assumed European superiority as a given, and the mere presence of a large dentition automatically led to the conclusion that the associated brain was therefore smaller and that the position of the possessor in the world was consequently “lower.” None other than the eminent 19th century biologist and defender of Darwin, Thomas Henry Huxley, declared that “no rational man . . . believes that the average negro is the equal, still less the superior, of the average white man.” He went on to say that “it is simply incredible that . . . our prognathous relative . . . will be able to compete successfully with his bigger-brained and smaller-jawed rival, in a contest which is to be carried on by thoughts and not by bites” (Huxley 1865:561).
Before I go on to deal with tooth size and the circumstances that relate to size differences between human populations, I should first dispel the idea that large teeth entail small brains. There is absolutely no relationship between brain size and tooth size (Brace et al. 1991:37). As with other traits of adaptive value, the selective forces appropriate to the control of one are completely unrelated to the forces responsible for the control of the other. People do not chew with their brains or think with their teeth. Consequently, variation in one trait is completely unrelated to variation in the other. Second, there is absolutely nothing to sustain the idea that “other” people have smaller brains than Europeans. When brain size is corrected for body size, there is no demonstrable difference between any of the populations of the world (Jensen 1984:54-55; Brace et al. 1991:37).
If Huxley was wrong in his assumption that the African brain is significantly smaller than the European brain, he was quite correct in his recognition of the fact that Europeans have smaller jaws and teeth. Oddly enough, that “fact” had not been demonstrated by any systematic quantitative study before he produced his declaration, and very little attention has been devoted to the matter since that time. As it happens, the issue of tooth-size differences between human populations past and present has been largely ignored by the dental and anthropological sciences.
No living human population has teeth that are as large on the average as their predecessors in the Middle Pleistocene, so it is evident that dental reduction has taken place over the last 100,000 years or more in the lines that have led to all of the regional human inhabitants of the world. Reduction has amounted to at least 40-45% in those who live in the north temperate zone between Europe and the Middle East. This is the maximum amount of dental reduction visible in modern Homo sapiens, although it is equaled in spots in eastern Asia such as Hong Kong and the northern island of Hokkaido in Japan. The least amount of reduction is evident in Australia where it runs between 10 to 15% (Brace 1980). In sub-Saharan Africa, the amount of reduction in the groups in the Congo Basin and in West Africa with the largest jaws and teeth is around 25%, but it runs up to 40% in the Horn of East Africa.
The documented reductions in the dentition have all taken place as “archaic” human form — what I would call regional representatives of the Neanderthal Stage — becomes transformed into “modern” Homo sapiens (Brace l991b:170, 1994). We are not accustomed to thinking of the Neanderthals as cultural innovators, but the very real possibility exists that they may have been “the ones who pioneered the use of cooking as a regular means of preparing food” (Brace l991b:155). Survival in the north during the onset of glacial conditions depended on the control of fire. Among other things, there would have been no point in going to the trouble of bringing down a large Pleistocene prey animal if the thing would become unusable within a day or so by virtue of having frozen solid. “Obligatory cooking” (Brace 1994a,b), then, was a part of the cultural traditions of those who survived through the last two glacial maxima along the northwestern edge of human habitation in the Old World, and it has been suggested that the extensive hearth residues that characterized the Mousterian sites occupied by the Neanderthals were actually the remains of earth ovens in which they regularly thawed the products of the chase simply in order to make them edible (Brace and Montagu 1977:335-336; Brace 1978:214, 1979:545-546, 1992:16-17; Brace et al. 1987:713-714, 1991:46-51).
Cooking not only made it possible to eat what previously had been frozen, but it reduced the amount of chewing necessary to get anything so treated into swallowable consistency. The regular use of cooking, then, reduced the amount of mandatory chewing. A reduction in the amount of tooth use over a lifetime meant that teeth did not wear out quite so fast, and also that a person did not need quite such a large dentition to be able to survive for the otherwise expectable human span. Under such conditions, mutations affecting tooth size could accumulate without being automatically weeded out by selection, and, since most such mutations produce a reduction in size, the predictable long-term results would be dental diminution. Today, the largest number of the people with the smallest teeth in the world are those who live in that stretch that runs from the Middle East through to the Atlantic Coast of Western Europe — just that area where the cooking of food has been in continuous use for the longest period of time.
Figure 4 shows a crude picture of relative tooth size among the modern populations of the Old World. The reader will remember that it was in just that stretch running from the Middle East to the west and north that pigment reduction began earliest and has gone to its greatest extent. Pigment, however, is related to the intensity of ultraviolet radiation, and teeth are only related to the nature of the tasks required to process food. Of course, food is more likely to freeze in areas of limited solar radiation, so there is a degree to which the value of maintaining skin pigmentation and the value of maintaining a dentition that could cope with uncooked food covary to a certain extent. Some of this is apparent when Figure 4 is compared with Figure 3, but it is also apparent that there are areas where the traits are completely uncorrelated.
The evident value of cooking at northern latitudes meant that it spread into the northeastern areas more readily than it did to the south. Dental reduction then followed suit, and tooth size in Eastern Asia followed the same track that it had taken starting somewhat earlier in the northwestern parts of the Old World. Eventually, of course, it spread south because, although it was not necessary to thaw food in the tropics, the repeated application of heat to the ripening results of a previous kill could partially delay and counteract the noxious effects of the various micro-organisms that would otherwise render the decaying products of earlier hunting activities unfit for human consumption.
Cooking was adopted last in Australia, and dental reduction is least evident among the aborigines who were encountered by the Europeans who settled there over the last two centuries. Earth oven cookery had spread to Australia perhaps before the end of the Pleistocene, but, in any case, the kind of reduction in chewing stress that it represented got into Australia later than anywhere else in the world. Not only that, but it spread slowly south in the continent and never did get to Tasmania before the terminal Pleistocene rise in sea level turned it into an island. As a consequence, there is a tooth-size gradient in Australia running from the north down to the south where people had the largest-sized teeth of any living human beings. Even so, tooth size had begun to reduce from the Middle Pleistocene-sized levels that were displayed only 10,000 years ago (Brace 1980:147).
Some of that recent reduction may have been as a result of gene flow from areas to the north where reduction had already taken place, but some may have been because of the relaxation of selection that followed the adoption of cooking techniques. At the moment, it is not possible to say how much of which led to that recent reduction. In any case, the pigment cline is reversed and skin color in the south lightens in just those areas where tooth size gets to its maximum. This is just another instance of the general principal that separate traits will respond to the changes in the intensity of the selective force that is responsible for their individual manifestation, and that each will go its own way without regard to what any other trait is doing unless that other trait is responding to that self-same selective force.
Late in the overall picture of dental reduction, another event occurred which completely changed things as far as the teeth were concerned. This was the invention of pottery. Archaeological reputations have been made and lost as a result of the analysis of pottery, but few have noted what the presence of the humble pot meant to the inventors and users. It meant that teeth were no longer necessary at all. Of course, we dearly enjoy the process of chewing the culinary delights that we savor, and we tend to think that the toothless are less attractive than those who are dentally well-endowed, but as soon as pots became available it was possible to reduce nourishment to drinkable consistency. From that point on, the edentulous could survive as well as anybody else. Since selection for the maintenance of tooth size all but ceased at that point, one would predict that recent human tooth size reduction should have accelerated to its maximum. From the evidence available from the several places in the world where pottery had more-or-less independent beginnings, this indeed appears to be the case.
For various reasons, that post-Pleistocene change has been entirely overlooked in the anthropological world, and we still hear that somewhat self-satisfied statement that once “modern man” had emerged, maybe 35,000 years ago, all biological evolution ceased and the only subsequent changes were in the cultural realm (R. Leakey in Fisher 1983:145; Diamond 1990:26; Klein 1992:12). A quick perusal of the actual evidence, however, shows that this is simply not so. Not only have jaws and teeth undergone a change of up to 20% since that time, but there has been an equally clear-cut reduction in the indications for muscularity and robustness in the post-cranial skeleton as well. Cultural change certainly has been important. In fact, it was the reduction in environmentally imposed selective forces created by those very cultural developments that led to the changes visible in the human physique.
When other genetically controlled aspects of human variation are mapped, they also tend to show unrelated distributions that are as independent of each other as those shown in Figures 2, 3, and 4. For example, when aspects of the common variance in the expression of some 95 genes are plotted on a world map in terms of principal component loadings, the distributions of the first three components are completely independent of each other (Cavalli-Sforza et al. 1993). One such map will show the greatest intensity of loadings in Africa with decreasing effects strictly proportional to distance. The authors have chosen to regard this as indications of an actual migration out of Africa at an unknown date in the past with modern differences being proportional to the time of separation. Another such map plotting the next independent residuum of common variance shows the greatest intensity of loadings in South America with decreases proportional to distance from that spot. Again, this distribution is presumed to illustrate the results of migration. The same is assumed to be true for plots of the remaining independent residuals of common variance.
The problem with trying to invoke migration as the explanation for such gradients in the manifestations of genetic likeness is that, while it might work to explain an initial distribution, the traces of that pattern would be completely altered by just one later migration from a separate center. The independent and non-coincident patterns of genetic distributions can only be the results of the operations of unrelated selective force gradients and the further complications added by genetic drift. At the moment, it is impossible to say how much of which has contributed to the picture of independent regional manifestations of genetic difference shown by the separate principal component maps. The only thing we can say for certain is that migrations cannot possibly be invoked to explain the distributions observed, and that the pattern is completely compatible with what would be expected if separate and unrelated selective force gradients were involved.
Intelligence
Of all the traits that have played a role in ensuring the survival and success of Homo sapiens as a species, innate brain power clearly must be accorded a position of prime importance. Just as the order Primates as a whole is distinguished from the rest of the animal kingdom by a greater average degree of intelligence and a corresponding expansion of the organ from which that emanates — the brain — so humans transcend their Primate relatives in this regard by yet another order of magnitude (Donald 1991:98- 100). The human brain underwent a near threefold expansion from the essentially chimpanzee-sized brain of our Australopithecine ancestors 1.5 to 2.0 million years ago. It stands to reason, then, that this illustrates the results of the continuing effects of strong forces selecting for the increase in intelligence. One might also think it logical that intelligence in the human world would be distributed in a manner analogous to that of other traits under strong selective force control, i.e. not in terms of population boundaries but according to the pattern of distribution of the relevant selective force. As we shall see, this is probably not the case. There are unexpectedly difficult problems involved in assessing both the selective forces involved and the nature of the distribution of intelligence.
Unfortunately, “intelligence” is another of those words like “race” that people use with glib facility but which proves to be extremely hard to define. I am not going to offer my own definition because it would be just another in a list that is as long as the number of people involved in comparing themselves to others. At bottom, “intelligence” is a completely subjective category. It has no clearly identifiable reality like a pigment or a hemoglobin molecule, and it has no tangible measurable form like a tooth. Its existence as a category at all is an example of the creative capacity inherent in one of the most powerful tools at human disposal —.language. The words we use are analogues of a real world that is vastly more graded and complicated than our verbal rendition. Of course we gain much of our power to deal with the infinite complexities of that graded reality by reducing it to a finite and manageable linguistic analogue. However, the danger always exists that our linguistic simplifications may just create categories where none really exist. “Race” clearly is an example of just that. “Intelligence” comes close to being another.
One of the problems stems from the essentially subjective way in which we perceive the phenomenon. Virtually all living human beings compare themselves to the people with whom they interact on what they perceive as a scale of relative cleverness. We all are perfectly convinced that we can assess relative intellectual ability, and each of us uses our own capacities as the basic yardstick. Consequently there are as many standards as there are individuals involved in the debate. Nor have the professional scholars involved in the assessment of intelligence provided much assistance. Possibly the most opaque and unhelpful non-description ever offered was the one coined by the distinguished psychologist, Edwin G. Boring: “Intelligence is what the tests test” (Boring 1923:35). That simply expands the possible scales of measurement beyond the number of individuals doing the evaluating by the amount of tests that professional ingenuity can concoct.
It has long been realized that the increase in brain size during the evolution of the mammals was certainly related to an increase in intelligence (Radinsky 1967, 1979). In parallel fashion, the increase in human brain size from the Pliocene through the Middle Pleistocene was also evidence of the increase in intelligence (Van Valen 1974). Relative brain size between living species also is associated with degrees of intelligence (Brace and Brace 1976). With this as a general background, there has been a repeated attempt to associate differences in brain size between the living human populations with differences in their intellectual abilities.
Before the American Civil War, the claim was made that the institution of slavery was justifiable on the basis of supposed differences in brain size and intelligence between African-Americans and European Americans (Nott 1849:35-36, 1858:77; Nott and Gliddon 1854). There are still psychologists who maintain that “among humans, crude brain size does have some relation to intelligence” (Rushton 1984:12), and that the best way to determine a persons intelligence is to “take a tape measure, put it around people’s heads, (and) measure the head circumference” (Rushton 1989).
The thing that is left out of these attempts to assess the relative brain power of living members of the same species is any correction for body size. From my own measurements of the crania of samples representing all of the major living populations of the world, I can show that the average difference between the largest and smallest heads is not significantly different from the average difference between male and female brain case size. In the latter instance in any given population, the male/female size difference is entirely in proportion to the difference in body size.
If one is going to argue that larger head size within a single species such as Homo sapiens really indicates greater intellectual power, then one will also have to support the proposition that, on the average, men are smarter than women. There is absolutely nothing to sustain such a conclusion, and the suspicion arises that those who are continuing to pursue research relating to the average “racial” differences in brain size are simply engaging in an exercise in applied bigotry.
Conscientious scholars in the intelligence-evaluation business have devoted much effort and ingenuity to the process of making intelligence tests “culture-free” or “fair” in a variety of ways, and one such has written a ponderous tome filled with numbers and formulae called Bias in Mental Testing (Jensen 1980). The basic assumption behind that work is that bias can be eliminated by proper procedural means. Beyond the subjectivity inherent in the assessment of individuals, however, there is the added problem associated with evaluations projected upon whole groups.. Professionals involved in the testing of intelligence simply accept the biological reality of “race” in the first place and go on to presume that it is reasonable that “racial” differences in intelligence are simply there to be discovered (Jensen 1969a:80, 1969b:14; Neary 1970:62). This is nothing less than bias itself, and no amount of statistical manipulation can eliminate it. Although it is presented with all the trappings of serious scientific scholarship, it is just a gussied-up version of the universal ethnocentric assumption held by every living human society that “we” are better than “they.”
In equally simplistic fashion, the further assumption has been gratuitously added that intelligence is the ability to adapt to “civilization,” and that “races” differ in intellectual ability in conjunction with the civilizations with which they are associated. This was one of the undocumented items of faith at the core of the Eugenics movement early in the century (Popenoe and Johnson 1918:285, 292), and it was shared with equally unjustified enthusiasm by the contemporary supporters of geographic determinism (Huntington 1915, 1924:1). One recent advocate of such a view feels that the Stanford-Binet I.Q. test measures the inherent ability to adapt to “Western civilization” (Jensen 1969b:14).
Nothing could better illustrate the dangers of making judgments about the relative worth of whole populations in the absence of the basic facts than potentially harmful and certainly irresponsible claims such as that. As we have seen above, even a relatively simple and clearly adaptive trait such as skin color shows only minimal response to alterations in selective force intensity that have been in effect for no more than 10,000 to 15,000 years. Our vaunted “Western civilization” has a time depth of only a small fraction of that span, and most of those who now reap its benefits are descended from ancestors who, only a few centuries back, were peasant farmers untrained in the niceties of reading and writing.
Even more to the point, there is not a single society in the world today that pursues a way of life like that of its ancestors 10,000 years ago. Even the few groups who lived by hunting and gathering until quite recently have all acquired steel tips for their spears and arrows, steel knives and axes, plastic containers, synthetic fiber cordage and firearms. Given the pace at which selection works on something as elusively complex as intelligence, a much more plausible case could be made for the view that virtually no living population is mentally adapted to the society with which it is now associated.
Now turn things around and look at them from the perspective of the conditions under which our mental capacities actually were shaped. Ten-thousand years ago, agriculture had yet to be invented. There were no sedentary farming communities, and subsistence was gained by foraging for what could be acquired by hunting and gathering activities. From that point on back for the previous million to 1.5 million years, hunting and gathering was the common heritage shared by all human beings. Surely it was that long-term situation which played the major controlling role in shaping human intellectual capabilities..
Of course, objections have been raised that the environment and the availability of specific resources was radically different in the disparate parts of the world. The geographic determinists of two or three generations ago were fond of contrasting the supposedly indolent life in the tropics where goodies were available for the plucking from a plethora of fruiting trees and bushes with what was purported to be the bracing rigor of more northerly climes where survival was said to depend more on ingenuity and disciplined effort.
Aside from the lesser chance of freezing to death, however, there is virtually no evidence to suggest that it is any easier to gain a living by gleaning from the land in the tropics than it is in the temperate portions of the world. The knowledge of what is edible and what is not, what ripens where at which time of the year, and the detailed habits of potential prey animals is every bit as difficult to come by at the equator as in the north. Wherever professional scientists have had extensive contacts with local populations who live off the land — whether it be Eskimos in the arctic, Australian aborigines in the “outback,” or the inhabitants of the New Guinea highlands barely south of the equator — they have come away with the realization that the people they have gotten to know all understand their local natural history with a sophistication and detail that is fully the equivalent of what would be required of a doctoral candidate defending a Ph.D. dissertation on aspects of the area in question. In the pre-literate world of the hunter-gatherer, evidently the penalty for stupidity is starvation.
Nothing can illustrate this point more graphically than the saga of the elderly Australian who led his group on a six-month trek to escape the consequences of the drought of 1943 in the outback of Western Australia. His first goal was the fallback waterhole at the extreme northwestern corner of the tribal territory which he had visited only once in his youth more than half a century earlier. When the resources there started to fail, he led them off westwards again through territory known to him only through the verses of a song cycle sung at totemic ceremonies and depicting the legendary wanderings of “ancestral beings.” The trek led on through a sequence of more than 50 waterholes until the little band finally emerged at Mandora Station on the coast of Western Australia more than 600 kilometers from where they had started (Birdsell 1979:147-148). If he had been wrong just once in his sequence, that would have been it for the whole group. Evidently the “myths” that constituted those tribal ceremonies actually represented the transmitted knowledge of previous generations, and, as the story shows, this could literally be of life or death significance. One would be hard put to come up with any instance in the literate world where survival was so directly dependent upon such a feat of human memory.
In their reliance on a major hunting element in their subsistence activities, ancestral humans depended on a most un-Primate-like kind of behavior. Even for those other Primates that occasionally catch and eat animal prey, there are none in which this kind of behavior is essential to their very survival. Prehistoric human beings, on the other hand, clearly relied on the products of the chase for their survival. And yet our basic anatomy gives almost no hint of a capacity for successful hunting activities. There is not a trace of specialization towards effective capture and disabling of prey to be found in the shape and the size of our teeth, and human locomotor capabilities are such that we cannot outrun anything larger than a rabbit.
Finally, human beings exploit a spectrum of plant foods that is orders of magnitude beyond the list relied on by even our cleverest non-human relatives. A fair portion of these would be of no value unless subjected to extensive preparation prior to ingestion. Human beings are the only creatures that cook their food, but, beyond that, no other animal has made more than the most rudimentary start at the ingenious roster of peeling, shelling, pounding, grating, grinding, soaking, leaching and the many other techniques that people have used to render some of the most unlikely items capable of yielding nourishment.
Surely it is these accomplishments and the capabilities that made them possible that are behind the successful spread of humans into such an extraordinary variety of locales. The ingenuity necessary to extract sustenance in the most unlikely areas and by the most unlikely means was an essential element in the human success story. That ingenuity was certainly taxed by the need to maintain a network of supportive relationships and deal with potential human competitors. It had to have been a combination of these factors that constituted the selective pressures that led to the expansion of the human brain starting at the beginning of the Pleistocene two-million years ago.
These circumstances, however, were a constant for human beings pursuing a hunting and gathering mode of subsistence at any time during the Pleistocene and throughout the length and breadth of their occupation. With the nature of selection for intellectual development held constant, the response should have been a constant also. Under such circumstances, there should have been no differential response. This is why human intelligence, although an adaptive trait of utmost importance, should not be expected to have a clinal distribution. If there is one faint exception to the expectation that there should be no significant difference between the intellectual capabilities of any of the human populations living in the world today, it would be of a most unexpected nature. It is a common prejudice of representatives of groups with long and sophisticated written traditions to regard their non-literate contemporaries with a kind of self-satisfied contempt. The denigrations run from regarding such people as “barbarians” or “benighted savages” to evaluations that barely consider them human. Almost invariably, their manifestations of what is considered to be cultural inferiority are assumed to be innate, and they are consigned to a state of genetically determined intellectual inferiority.
On the other hand, as the previously mentioned Australian saga abundantly demonstrated, it surely requires more basic smarts to survive in a world where one cannot go and look up the answers to crucial questions in a book. For those of us who can consult reference works in a library, it is just less important that we keep that information stored in our heads. It is just possible, then, that those whose ancestry has the longest continuous tie with literate civilization are the recipients of a heritage where selection for intelligence has been least stringent. Now recall that the rationale of the Probable Mutation Effect suggests that where the strength of selection maintaining a given trait is relaxed, there should be a consequent reduction in that trait itself. If indeed there is any difference in intelligence among the living populations of the world, then the greatest innate wit should be found among those who have most recently emerged from a non-literate past.
No representatives of a tradition of continuous literacy, however, have been able to enjoy its benefits for much more than 5,000 years at the most. As we have been able to see, tooth size may have changed as much as 5% during that time, but that figure is so close to measurement error that we cannot take it as proof of real biological difference. And no discernible skin color difference has arisen during that time even though some populations have moved into areas where selection operating over much longer periods of time has produced highly visible consequences. Therefore, even though the PME suggests that there might be a slight difference in intellectual capability between living human groups in the opposite direction from that so commonly assumed, the problems associated with assessing the amount of innate intelligence both within and between human populations are so sticky that it is unlikely that we shall ever be able to put such a
possibility to the test. At the moment, we have no reason to suspect that there are any group-specific differences in intelligence, and we have no way to test for them even if we did.
Clusters
There are other traits that are distributed according to the graded effects of the selective forces that influence their appearance. The relative length of the lower arm and leg in proportion to the upper arm and leg, and limb length in proportion to trunk length are related to conditions of heat and cold. Average length and elevation of the nose on its part is related to the relative lack of moisture in the air. With the sole exception of intelligence which is equally important everywhere and consequently everywhere the same, it would seem that every trait that is really important for human survival is distributed in conjunction with the intensity of the controlling selective force, and that these on their part are distributed completely without regard for human population boundaries.
From this it follows that the traits which are distributed across population boundaries as though they did not exist are the only ones that have real biological significance, while those that are constrained by population boundaries have no differential survival value. Although the more orthodox of R. A. Fisher’s followers still feel that we are obliged to assume that every biological trait is determined by a controlling selective force even if we cannot figure out what that might be, others are increasingly coming to the realization that there are many configurations that have no particular adaptive value in and of themselves. Those who have claimed that the failure to find adaptive meaning in certain traits is simply due to limitations in our powers of understanding have been called “hyperselectionists” and compared to “foolish Dr. Pangloss” of Voltaire’s 18th century satire, Candide (Gould and Lewontin 1979; Gould 1980:40). In Dr. Pangloss’ philosophy, “all is for the best in this the best of all possible worlds,” even if it is beyond our capacity to understand what might be good about it.
As disaster after disaster befell the unfortunate Candide, it became quite apparent that there was not always some unappreciated “good” behind the events of the world. In like fashion, it is evident that many biological configurations are that way just because of chance circumstances and not because of any unappreciated benefit conferred on their possessors. This realization had been reinforced by the discovery that there are substantial sequences of DNA in the genome that are just there and serve no functional purpose — “junk DNA” in the words of one geneticist (Ohno 1972).
In the absence of selective force control, certain variants may increase or decrease in frequency in various regions simply by chance. The process referred to by Sewall Wright as “genetic drift” can produce local differences in inherited features particularly where the populations are relatively small and partially isolated from their neighbors -circumstances that were generally the rule for the human condition prior to the advent of agriculture (Wright 1943). The significant expansion in numbers by those groups that adopted a farming mode of subsistence subsequently established certain of those chance local variants as the characteristic regional morphological conformation in the various areas where agriculture had led to that notable expansion in population size. The ensuing perpetuation of identifiable regional configurations then occurred for the simple reason that the pattern was already present — not because it conveyed any particular adaptive advantage.
We can identify many such aspects of morphology associated with the configuration of the heads and faces of people in various parts of the world. Some of these coexist with features that are legitimately under selective force control, and we have to make an effort to separate the two categories. For example, nasal form in the long-term inhabitants of the colder portions of the world runs to the high-bridged and elongate. The Native Americans in the northernmost portions of the New World were famous for having noses that were just as prominent as those of northern Europeans. However, in the other details of the face — for instance, the jut of the cheekbones and the relative forward placement of the lateral borders of the eye sockets — they were quite distinct from characteristic European form although they could not be differentiated from the positioning of cheekbones and lateral orbital margins of the living people of northeast Asia.
Nasal form is adaptive and responds to the forces of selection, but there is nothing particularly adaptive about nuances of cheekbone form or shape of the eye socket. What possible advantage could it be to have high rounded eye sockets as opposed to low wide rectangular ones? (Brace and Hunt 1990:344). And why would it be better to have cheekbones that are hollowed out below the lower margin of the orbit as opposed to ones that drop straight down to present the appearance of a relatively flat face? Differences in the details of facial form of this nature are clearly associated with different regions of the world, but it is hard to see the faintest reason why any one should be any “better” than any other. And yet each region shows a continuity running from the present back into the past to the point where general morphology ceases to look “modern.” In fact, some of those very aspects of cheekbone and eye socket morphology that show different configurations in different regions of the world today were also characteristic of facial form for the archaic inhabitants of the same parts of the world going back into the Middle Pleistocene.
The same sort of thing is true for the form of the rest of the skull. For example, the characteristic general cranial shape at the northwest edge of Europe is long and low with a tendency towards the presence of a marked bulge or “bun” at the rear, while the characteristic form in eastern Europe is towards a skull that is short, wide and almost flat behind (Brace l991a). But, again, there is absolutely no reason why one such shape is “better” than another, and, despite undocumented claims to the contrary (Cavalli-Sforza et al. 1993:641), no possible set of selective forces can be imagined that would have led to the special development of either one. It really does not matter what shape the brain case takes so long as it is large enough to encompass the brain. At best, we are left to assume that the different local patterns were simply the products of genetic drift (Lynch 1989:13). Once established, these are perpetuated because that is what the local gene pool has to contribute to subsequent generations.
What is true for so many aspects of the bones of the head is also the case for many of the details of its fleshy covering. The external shape of the eye opening differs from one human population to another. For example, it is widely recognized that people of eastern Asian ancestry are likely to display a particular and recognizable configuration. However, there is no evidence that it has any effect whatsoever on the ability to see. At one time, it was suggested that the people of eastern Asia had been shaped by prolonged exposure to intense cold during the last glaciation, and that this led to the developments of eyelid form that played a protective role (Coon et al. 1950:71). However, the ancestors of Europeans survived the effects of the last two glaciations without ever developing the eyelid form characteristic of modern eastern Asians, and there is no indication that the characteristic form of the European eye opening puts them at any disadvantage even when subject to prolonged exposure to conditions of intense cold. The form of the external ear also differs in characteristic fashion from region to region. But again, there is virtually no evidence to suggest that there is an associated difference in the ability to hear or that selection is involved. And the same thing can be said for the fleshy portions of the lips and nose.
It is by an appraisal of features such as these that we can recognize the portion of the world to which the ancestors of a given person are likely to have belonged. When we take a standard set of measurements on representatives of people from all parts of the world, the different samples will show patterns of regional association. All the samples from a given region will cluster with each other as opposed to samples from another region unless, of course, a given sample actually is composed of migrants from somewhere else. When that is the case, the migrant sample will cluster with samples from the region from which it originally came.
In an effort to create a picture of clusters for all of the major regions of the world, I have made measurements on crania from as many samples as I could get access to over the last dozen years. These were then converted into standard scores as Howells recommended (Howells 1986). Sample means were compared by an unweighted pair grouping method analysis (UPGMA) and used to create a Euclidean Distance dendrogram (Romesburg 1984:15; Brace and Hunt 1990:347). This is the same procedure that was used to generate the pattern shown in Figure 1.
When samples from all over the world were treated in this fashion, the picture that emerged is that seen in Figure 5. At that, it is greatly simplified from the much larger cluster diagram that was generated when I used more than twice as many groups as are depicted in Figure 5. Even so, it is obvious that the samples from each region tend to cluster with each other. When the various samples from each region were merged with their nearest neighbors so that only a single pooled group was left and a Euclidean Distance dendrogram was produced, what emerged was the picture seen in Figure 6 In this, the individual twigs stand for the regional clusters from which they were formed and can be called by the name of the region with which it is associated.
So we can refer to an African Cluster, an East Asian Cluster, a European Cluster and so forth. It would be a mistake to say that this is just another name for “race,” since nothing held in common by the members of any cluster allows us to say anything about the adaptations of its components. The only thing that ties members of a given cluster together is the common inheritance of traits of no adaptive value whatsoever. A cluster by its very nature cannot be assessed in terms of ability, intelligence, resistance to disease or anything else that could be placed on a hierarchy of value since the only thing that holds a cluster together in an analytical sense is the complete triviality of the traits by which it is defined.
Nor is there any significance to the mean figures calculated by generating the central tendencies of the clusters as is done to construct Figure 6. It is important to note that those “central tendencies” do not reflect any kind of “essence” or aspect of common variance. The farther away from the center of a geographic region one goes, the less the peripheral members of that cluster share with the others in an opposite direction and the more they share with members of the immediately adjacent geographic region. Major regional cluster names, then, are simply convenient labels for describing people who differ in utterly unimportant ways. The convenience offered by geographic names is that there is no possible loading with the hint of the often covert implications included when descriptive or ethnic labels are used. Finally, geographic names can be easily amended by directional modifiers. Asia can be sharpened by specifying Central Asia, Western Asia, South Asia, Southeast Asia and the like. The same is also true for the other major clusters such as Africa, Amerind, Australia, Europe, Pacific-Ainu and Siberia-Eskimo.
Conclusions
Where Homo sapiens is concerned, the concept of “race” does not correspond to any biological entity. Not only does it have at least as many flaws as the concept of subspecies (Wilson and Brown 1953; Brown and Wilson 1954), but it also has been invoked as a platform for the purpose of sustaining invidious value judgments that stem largely from unwarranted prejudice. Its existence arises entirely from the human tendency to categorize when dealing with what is perceived to be the nature of the world. In this case, the category “race” is a misperception of the reality of human biological variation.
That variation exists, and it is real and significant. However, those aspects that have arisen in response to the effects of natural selection cannot be perceived or understood if we take popularly assumed “races” as our starting points. Only by tracing the distribution of each trait separately can we discern its association with the relevant selective force. Since selective forces themselves have distributions that are unrelated to human population boundaries, traits that are under selective force control, and hence that are biologically significant, will also be distributed without relation to those human population boundaries. More than one modern anthropologist has clearly perceived these matters and treated them with admirable sophistication (Bermann 1992; Sauer 1992).
Where traits are constrained by human population boundaries, the dynamics by which they are controlled are entirely the result of inheritance and local breeding behavior. The manifestation and distribution of such traits is unrelated to the forces of natural selection and therefore will have no adaptive value. As a consequence, they cannot be ranked as “better” or “worse” when populations are compared with each other. At most, they can be seen as “different” and as illustrating what has been perpetuated by chance in the different areas of human habitation.
Regional configurations observed in the clusters of samples from given parts of the world cannot be labeled by using aspects of variation that are under selective force control. Designations such as “Negroid,” for example, fail to specify people from a given circumscribed area. Ethnic designations are no better. “Mongoloid,” for instance, is a word derived from the Mongols who are both geographically and morphologically peripheral to the other populations with whom they are tied. “Caucasoid” refers to the people living in the locale where, according to the Biblical narrative, Noah’s Ark disgorged its contents after the retreat of the Scriptural flood, and its use in a broader context implies acceptance of a whole series of assumptions that have no scientific basis.
The only really useful means of referring to the various regional configurations visible in the world is by using broad geographical terms to designate the appropriate inhabitants. The people of Europe are properly referred to as Europeans, the people of Africa are Africans, the people of Australia are Australians, and so forth. This can be narrowed down by referring to people as South Asians, Northwest Europeans, Southwest Australians and the like. That way no “ethnic” or behavioral attributes are indicated, and no confusion or hidden implications can result from the labels given.
Finally, if indeed that all-important but hopelessly undefinable phenomenon, “intelligence,” has the adaptive significance for human survival that we have reason to assume it does, then it has to be apportioned according to the distribution of its controlling selective force. Further, if the latter is distributed in a manner analogous to that of other forces affecting human survival, it is clear that we have no right to base our expectations concerning intellectual capability on a distribution that is limited by the boundaries marking the area of extent of any given population. Beyond that, no human population now lives under anything like the kind of circumstances that shaped human capabilities through that million-years-plus stretch of the Pleistocene during which they were established.
However, until 10,000 years ago or less, all humans everywhere were pursuing the same kind of subsistence strategy, whatever the latitude. This meant that the selective pressures influencing intellectual development at any given time were identical for all living humans over a span of more than a million years. Only within the last 10,000 years have things changed to the extent that it requires substantially more — or less — intelligence to make a go of it in some parts of the world as opposed to others. That relatively short post-Pleistocene stretch is not enough to produce a measurable difference between the intellectual capacities of the various surviving populations of the world.
Given all of this, we have no right to expect that there is any innate difference of intelligence between the various regional clusters and configurations of modern human beings. The only possible reason behind efforts to test for differences in “racial” intelligence is an assumption — whether covert or overt — that some groups are brighter and therefore better than others and that one’s own group can be shown to be the brightest and best. Since the configurations present in regional clusters are nothing more than ‘family resemblance writ large’ and have no other biological meanings, any attempt to assess “racial” difference in any aspect at all with the expectation of finding something of significance beyond an otiose catalogue of trivia can only be based on invidious expectations that themselves can have no scientific justification.
“Race” at best is just another four-letter word, empty of any biological significance. Biological differences do exist among modern humans, and an investigation of these differences is interesting and valuable. But this can only be accomplished by abandoning the “race” concept and tracing the traits to be studied according to their own distributions as they reflect the selective forces that have influenced the variation observed. Insofar as the assumption remains that regionally circumscribed peoples possess an essential nature defined by associated abilities, this is just a continuing manifestation of the use of the concept of “race” in the absence of the use of the name. The continuation of such an outlook can only result in scientific futility and social harm. As a concept without redeeming social or scientific value, “race” indeed deserves to be ranked with the roster of those four-letter words that merits the designation obscenity. It is too much to expect that what I say here will have much influence on the public at large, but at least I hope that it will be read and understood by the anthropological community and that they will respond by consigning the term and the concept to the oblivion that it deserves.
Rudyard Kipling, known as a jingoistic apologist for the British Empire, also had another side that reflected a deeper understanding of and sympathy for the essential humanity of the various manifestations of the “other” with which he had come in contact. Some of this is expressed in the conclusion of the verse that stands at the head of my Introduction, and it can serve as a fitting note on which to conclude:
But if you live over the sea,
Instead of over the way,
You may end by (think of it!) looking on We
As only a sort of They!
Acknowledgments
This manuscript is condensed from parts of a book in preparation entitled Race is a Four-Letter Word. Like the present paper, that book, when completed, is dedicated to Ashley Montagu, not only in honor of the splendid example he has set for us in an extraordinary lifetime of accomplishments motivated by the highest and most admirable of ideals, but also for the kindness and support that he has shown to me personally. I also wish to express my gratitude to Professors Larry J. Reynolds and Leonard Lieberman for giving me the opportunity of being part of this tribute to Ashley Montagu. The world is a better place because of him, and he continues to be an inspiration to us all.
Figure 1.
A Euclidean Distance dendrogram or cluster diagram demonstrating the atypical status of Mongols when compared with representative samples of the other inhabitants of East Asia. The values for the measurements were converted into C scores and compared by Unweighted Matched Pair Group Analysis (IJMPGA). The procedure was pioneered by Howells (1986) and is described in greater detail in Brace and Hunt (1990).
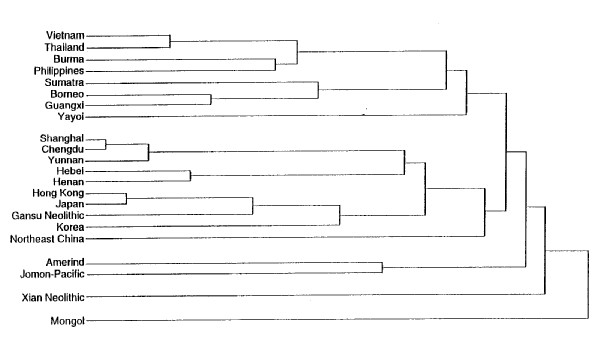
Figure 2.
The distribution of hemoglobin S according to its different frequencies in the populations of the Old World. Adapted from Bodmer and Cavalli-Sforza (1976:310). The various shadings represent the different indicated gene frequencies for hemoglobin S.
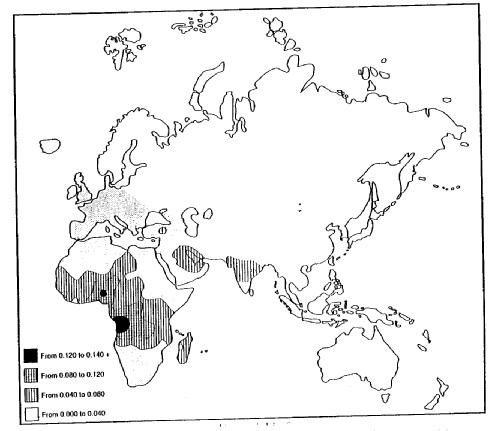
Figure 3.
The distribution of the variation in intensity of human skin color in the Old World. Adapted from Biasutti (1959:I:Table IV [pp.192-193]) by Brace and Montagu (1977:392), revised from Brace 1964:109).
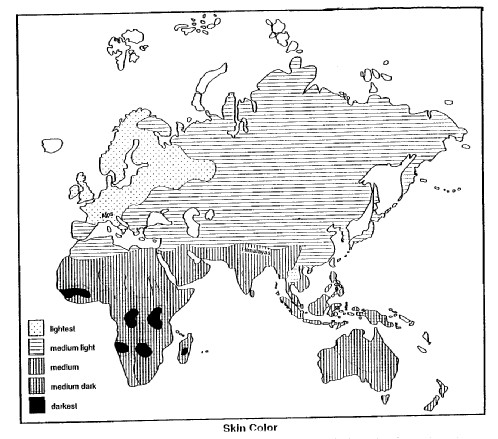
Figure 4.
The distribution of human tooth size among the populations of the Old World. A sampling of those data is reported in Brace et al. (1991).
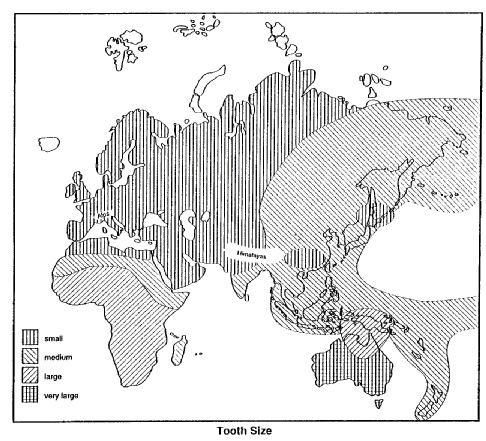
Figure 5.
A cluster diagram showing the relationships of samplings from the major regions of the world. The measurements used, and the samples are reported in Brace and Hunt (1990) and Brace and Tracer (1992). Variable transformation procedures were those used in Li et al. (1991) and Brace and Tracer (1992), while the cluster algorithm is the same as that used to generate Figure 1.
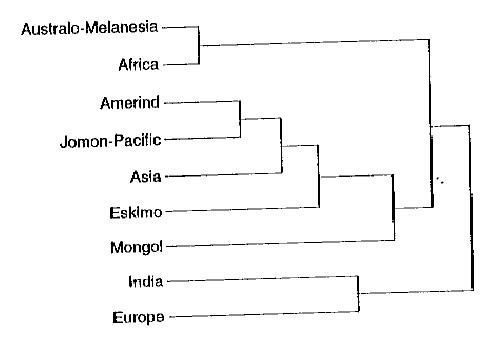
Figure 6.
A cluster diagram constructed by merging samples of each region to generate separate branches representing the cluster associated with each of the major geographic regions of the world. The procedure is the same as that used to generate Figures 1 and 5. The Mongols were left in, not because they represent a “major geographic region,” but simply to reinforce the point that they cannot be used to typify the inhabitants of eastern Asia.
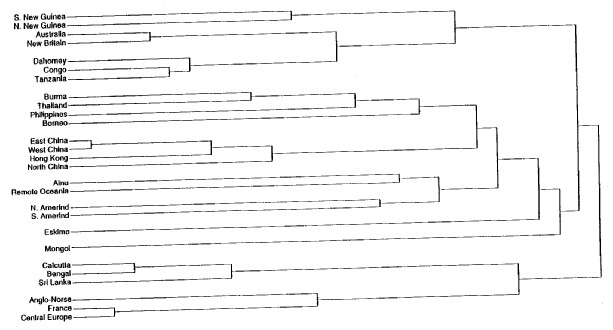
References Cited
Ackerman, L.V., J.A. del Regato, H.J. Spjut, and J.D. Cox 1985 Ackerman and del Regato’s Cancer: Diagnosis, Treatment and Prognosis. 6th edition. St. Louis: Mosby.
Barth, Frederick 1953 The southern Mongoloid migration. Man 52:5-8.
Bellwood, Peter S. 1979 Man’s Conquest of the Pacific: The Prehistory of Southeast Asia and Oceania. Oxford: Oxford University Press.
———. 1986 Prehistory of the Indo-Malayan Archipelago. Orlando: Academic.
———. 1991 The Austronesian dispersal and the origin of languages. Scientific American 265:88-93.
Benedict, Paul K. 1942 Thai, Kadai and Indonesian. American Anthropologist 44:576-601.
Bermann, Marc 1992 European exploration and the growth of racialist thought. Manuscript, files of author, Department of Anthropology, University of Pittsburgh.
Biasutti, Renato 1959 Le Razze e Popoli Della Terra: Volume I, Razze, Popoli e Culture. 3rd edition. Torino: Unione Tipografico-Editrice, Torinese.
Birdsell, Joseph B. 1979 Ecological influences on Australian Aboriginal social organization. In Primate Ecology and Human Origins: Ecological Influences on Social Organization. Irwin S. Bernstein Euclid O. Smith, eds. pp. 117-151. New York: Garland.
Blum, Harold F. 1959 Carcinogenesis by Ultraviolet Light: An Essay in Quantitative Biology. Princeton: Princeton University Press.
Blumenbach, Johann Friedrich 1865 The Anthropological Treatises of Johann Friedrich Blumenbach. Thomas Bendyshe, ed. 406 pp. London: Longman, Green, Longman, Roberts and Green.
———. 1969 De Generis Humani Varietate Nativa (1795). On the Natural Varieties of Mankind. Thomas Bendyshe, ed. and trans. New York: Bergman Publishers.
Bodmer, Walter F., and L. L. Cavalli-Sforza 1976 Genetics, Evolution and Man. San Francisco: Freeman.
Boring, Edwin G. 1923 Intelligence as the tests test it. New Republic 35:35- 7.
Box, Joan Fisher 1978 R.A. Fisher: The Life of a Scientist. New York: Wiley.
Brace, C. Loring 1963 Structural reduction in evolution. The American Naturalist 7:39-49.
———. 1964 A non-racial approach toward the understanding of human diversity. In The Concept of Race. M.F. Ashley Montagu, ed. pp. 103-152.
———. 1978 Tooth reduction in the Orient. Asian Perspectives 19:203-219.
———. 1979 Krapina, “classic” Neanderthals, and the evolution of the European face. Journal of Human Evolution 8:527550.
———. 1980 Australian tooth-size clines and the death of a stereotype. Current Anthropology 21:141-164.
———. 1991a Monte Circeo, Neanderthals, and continuity in European cranial morphology, a rear end view. In The Circeo I Neandertal Skull: Studies and Documentation. Marcello Piperno and Giovanni Scichilone, eds. pp. 175- 95. Rome: Istituto Poligrafico e Zecca Dello Stato, Libreria Dello Stato.
——— l991b The Stages of Human Evolution. 4th edition. Englewood Cliffs, New Jersey: Prentice-Hall.
———. 1992 Modern Human Origins: Narrow Focus or Broad Spectrum? The David Skomp Lecture. Bloomington, Indiana: Department of Anthropology, Indiana University. 30 pp.
———. 1994 Modern human origins and the dynamics of regional continuity. In Prehistoric Mongoloid Dispersals. T. Akazawa and E. Szathmary, eds. New York: Oxford University Press. In press a
———. 1994 Trends in the evolution of human tooth size. In Aspects of Dental Biology: Palaeontology, Anthropology and Evolution. Jacopo Moggi-Cecchi, ed. pp. 6-14. Florence: Angelo Pontecorboli Editore.
Brace, C. Loring, and Mary L. Brace 1976 Monkey business and bird brains. In The Measures of Man. Eugene Giles and Jonathan S. Friedlaender, eds. pp. 54-71. Cambridge, Massachusetts: Peabody Museum Press.
Brace, C. Loring, and Kevin D. Hunt 1990 A non-racial craniofacial perspective on human variation: A(ustralia) to Z(uni). American Journal of Physical Anthropology 88:341-360.
Brace, C. Loring, and Ashley Montagu 1977 Human Evolution: An Introduction to Biological Anthropology. 2nd edition. New York: Macmillan.
Brace, C. Loring, Karen R. Rosenberg, and Kevin D. Hunt 1987 Gradual change in human tooth size in the late Pleistocene and post-Pleistocene. Evolution 41:705-720.
Brace, C. Loring, Shelley L. Smith, and Kevin D. Hunt 1991 What big teeth you had Grandma! Human tooth size past and present. In Advances in Dental Anthropology. Marc A. Kelley and Clark Spencer Larsen, eds. pp. 33-57. New York: Wiley-Liss.
Brace, C. Loring, and David P. Tracer 1992 Craniofacial continuity and change: A comparison of Late Pleistocene and Recent Europe and Asia. In The Evolution and Dispersal of Modern Humans in Asia. Takeru Akazawa, K. Aoki, and T. Kimura, eds. pp. 439- 471. Tokyo: Hokusen-sha.
Brown, William L., Jr., and Edward O. Wilson 1954 Points of View. The case against the trinomen. Systematic Zoology 3: 174- 176.
Bulbeck, David 1982 A re-evaluation of possible evolutionary processes in Southeast Asia since the late Pleistocene. Bulletin of the Indo-Pacific Prehistory Association 3: 1-21.
Cavalli-Sforza, Luigi L., Paolo Menozzi, and Alberto Piazza 1993 Demic expansions and human evolution. Science 259:639-646.
Coon, Carleton S. 1939 The Races of Europe. New York: Macmillan.
Coon, Carleton S., Stanley M. Garn and Joseph B. Birdsell 1950 Races: A Study of the Problems of Race Formation in Man. Springfield, Illinois: Charles C. Thomas.
Cuvier, Georges 1817 Le règne animal distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction à l’anatomie comparée. Volume I: L’introduction, les mammifères et les oiseaux. Paris: Deterville.
———. 1829 Le règne animal. 2nd edition. Paris: Deterville and Crochard.
Daniels, Farrington, Jr. 1969 Optics of the skin as related to ultraviolet radiation. In The Biologic Effects of Ultraviolet Radiation (With Emphasis on the Skin) . F. Urbach, ed. pp. 151-157. Oxford: Pergamon.
Daniels, Farrington, Jr., J.C. van der Leun, and B.E. Johnson 1968 Sunburn. Scientific American 219:38-46.
Darwin, Charles R. 1859 On the Origin of Species By Means of Natural Selection, Or, The Preservation of the Favoured Races in the Struggle for Life. London: John Murray.
Diamond, Jared 1990 A pox upon our genes. Natural History 2:26-30.
Donald, Merlin 1991 Origins of the Modern Mind: Three Stages in the Evolution of Culture and Cognition. Cambridge, Massachusetts: Harvard University Press.
Fisher, Helen E. 1983 Richard Leakey’s time machine. Interview. Omni 5:94-104, 142-145.
Fisher, R.A. 1930 The Genetical Theory of Natural Selection. Oxford: The Clarendon Press.
Fisher, R.A., and E.B. Ford 1950 The ‘Sewall Wright’ effect. Heredity 4:117- 119.
Friedman, Robert J., Darrel Rigel, Alfred Kopf, Matthew Harris, and Daniel Baker 1991 Cancer of the Skin. Philadelphia: Saunders.
Gould, Stephen Jay 1980 Wallace’s fatal flaw. Natural History 89:26-40.
Gould, Stephen Jay, and Richard C. Lewontin 1979 The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proceedings of the Royal Society of London B 205:581-598.
Greenberg, Joseph H., and Merritt Ruhlen 1992 Linguistic origins of native Americans. Scientific American 267:9499.
Haynes, C. Vance 1982 Were Clovis progenitors in Beringia? In Paleoecology of Beringia. David M. Hopkins, J.V. Matthews, Jr., C.E. Schweger and S.B. Young, eds. pp. 383-398. New York: Academic Press.
Hoffecker, John F., W. Roger Powers, and Ted Goebel 1993 The colonization of Beringia and the peopling of the New World. Science 259:46-53.
Holick, Michael E. 1987 Photosynthesis of vitamin D in the skin: effect of environmental and life-style variables. Federation of American Societies for Experimental Biology. Federation Proceedings 46:1876-1882.
Howells, William W. 1986 Physical anthropology of the prehistoric Japanese. In Windows on the Japanese Past: Studies in Archaeology and Prehistory. R.J. Pearson, G.L. Barnes, and K. Hutterer, eds. pp 85-99. Ann Arbor, Michigan: Center for Japanese Studies, University of Michigan.
Huntington, Ellsworth 1915 Civilization and Climate. New Haven, Connecticut: Yale University Press.
———. 1924 The Character of Races. New York: Scribner and Sons.
Huxley, T. H. 1865 Emancipation — black and white. The Reader. Saturday, May 20, pp. 561-562.
Jensen, Arthur R. 1969a How much can we boost IQ and scholastic achievement? Harvard Educational Review 39:1-123.
———. 1969b Edited reply to letter by D.N. Robinson. New York Times Magazine, September 21, p. 14.
———. 1980 Bias in Mental Testing. New York: The Free Press.
———. 1984 Sociobiology and differential psychology: The arduous climb from plausibility to proof. Annals of Theoretical Psychology 2:49-58.
Kipling, Rudyard 1990 Rudyard Kipling: The Complete Verse, with a foreword by M. M. Kaye. London: Kyle Cathie Limited.
Klein, Richard G. 1992 The archeology of modern human origins. Evolutionary Anthropology 1:5-14.
Kligman, Albert M. 1969 Comments on the stratum corneum. In The Biologic Effects of Ultraviolet Radiation (With Emphasis on the Skin) . F. Urbach, ed. pp. 165-167. Oxford, Pergamon.
Kligman, Lorraine H. 1986 Photodamage to dermal connective tissue. In The Biological Effects of UVA Radiation. F. Urbach and R.W. Gange, eds. pp. 98-103. New York: Praeger.
Li Yongyi, C. Loring Brace, Gao Qiang and David P. Tracer 1991 Dimensions of face in Asia in the perspective of geography and prehistory. American Journal of Physical Anthropology 85:269279.
Livingstone, Frank B. 1958 Anthropological implications of sickle cell gene distribution in West Africa. American Anthropologist 60:533-562.
———. 1962 On the non-existence of human races. Current Anthropology 3:279.
———. 1967 Abnormal Hemoglobins in Human Populations: A Summary and Interpretation. Chicago: Aldine.
———. 1985 Frequencies of Hemoglobin Variants: Thalassemia, the Glucose-6-phosphate Dehydrogenase Deficiency, G6PD Variants, and Ovalocytosis in Human Populations. New York: Oxford University Press.
———. 1989a Simulation of the diffusion of the B-globin variants in the Old World. Human Biology 61 :297-309.
———. 1989b Who gave whom hemoglobin S: The use of restriction site haplotype variation for the interpretation of the Beta s-globin gene. American Journal of Human Biology 3:289-302.
Lynch, Michael 1989 Phylogenetic hypotheses under the assumption of neutral quantitative-genetic variation. Evolution. 43:1-17.
Mercier, Norbert, H. Valladas, G. Valladas, and A. Jelinek 1992 New Tabun cave chronology: TL dating of burnt flints from the Jelinek’s excavation. Book of Abstracts for the 3rd International Congress on Human Paleontology. Journal of the Israel Prehistoric Society, Supplement 1, p. 80.
Montagu, Ashley 1942 Man’s Most Dangerous Myth: The Fallacy of Race. New York: Columbia University Press.
Morton, Samuel George 1839 Crania Americana; or, A Comparative View of the Skulls of Various Aboriginal Nations of North and South America; to which is prefixed An Essay on the Varieties of the Human Species. Philadelphia: J. Dobson.
Murray, Hugh 1834 An Encyclopaedia of Geography. London: Longman, Rees, Orme, Brown, Green, and Longman.
Myrdal, Gunnar 1944 An American Dilemma: The Negro Problem and Modern Democracy. New York: Harper.
Neary, John 1970 A scientist’s variations on a disturbing racial theme. Life 68:58B, 58C, 58D, 61, 62, 64, 65.
Nichols, Johanna 1990 Linguistic diversity and the first settlement of the New World. Language 66:475- 521.
Nott, Josiah Clark 1849 Two Lectures on the Connection Between the Biblical and Physical History of Man. Delivered by Invitation from the Chair of Political Economy, etc., of the Louisiana University in December, 1848. New York: Bartlett and Welford.
———. 1858 Influence of anatomy on the march of civilization. New Orleans Medical and Surgical Journal 15:64-77.
Nott, Josiah Clark, and George R. Gliddon, eds. 1854 Types of Mankind. Philadelphia: Lippincott, Grambo.
Ohno, Susumu 1972 So much “junk” DNA in our genome. In Evolution of Genetic Systems. H.H. Smith, ed. pp. 366- 70. New York: Gordon and Breach.
Ortiz de Montellano, Bernard R. 1991 Multicultural pseudoscience. Spreading scientific illiteracy among minorities. Skeptical Inquirer 6:46-50.
———. 1992 Avoiding Egyptocentric pseudoscience: Colleges must help set standards for schools. Chronicle of Higher Education 38:B 1-B2.
Pathak, Madhu A., and Kathleen Stratton 1969 Effects of ultraviolet and visible radiation and the production of free radicals in skin. In The Biologic Effects of Ultraviolet Radiation (With Emphasis on the Skin). F. Urbach, ed. pp. 207-222. Oxford: Pergamon.
Patterson, David 1987 The causes of Down syndrome. Scientific American 257:52-60.
Pauling, Linus 1949 Sickle cell anemia: A molecular disease. Science 10:64-66.
Pawley, Andrew, and Roger Green 1973 Dating the dispersal of the Oceanic languages. Oceanic Linguistics 12:167.
Pietrusewsky, Michael 1992 Japan, Asia and the Pacific: A multivariate craniometric investigation. In Japanese as a Member of the Asian and Pacific Populations. Kazuro Hanihara, ed. pp. 9-52. Kyoto: International Research Center for Japanese Studies, International Symposium 4.
Pietrusewsky, Michael, Li Yongyi, Shao Xiangqing, and Nguyen Quang Quyen 1992 Modern and near modern populations of Asia and the Pacific: A multivariate craniometric interpretation. In The Evolution and Dispersal of Modern Humans in Asia. T. Akazawa, K. Aoki, and T. Kimura, eds. pp. 531-558. Tokyo: Hokusen-sha.
Pope-Hennessy, James 1968 Sins of the Fathers: A Study of the Atlantic Slave Traders 1441-1807. New York: Knopf.
Popenoe, Paul Bowman, and Roswell Hill Johnson 1918 Applied Eugenics. New York: Macmillan.
Potten, Christopher S. 1985 Radiation and Skin. Philadelphia: Taylor and Francis.
Radinsky, Leonard 1967 Relative brain size: a new measure. Science 55:836-838.
———. 1979 The Fossil Record of Primate Brain Evolution. The 49th James Arthur Lecture. New York: American Museum of Natural History.
Roberts, Richard G., Rhys Jones, and M.A. Smith 1990 Thermoluminescence dating of a 50,000-year old human occupation site in northern Australia. Nature 345:153-156.
Robins, Ashley H. 1991 Biological Perspectives on Human Pigmentation. New York: Cambridge University Press.
Romesburg, H. Charles 1984 Cluster Analysis for Researchers. Belmont, California: Lifetime Learning Publications.
Rushton, J. Philippe 1984 Sociobiology: toward a theory of individual and group differences in personality and social behavior. Annals of Theoretical Psychology 2:1-48.
———. 1989 Statement made on the CBC radio program, Quirks and Quarks, February 18.
Sauer, Norman J. 1992 Forensic anthropology and the concept of race: If races don’t exist, why are forensic anthropologists so good at identifying them? Social Science and Medicine 4:107-111.
Solheim, Wilhelm G., II 1981 Philippine prehistory. In The People and Art of the Philippines. Father Gabriel Casal, Regaldo Trota Jose, Jr., Eric S. Casino, George R. Ellis, and Wilhelm G. Solheim II, eds. pp. 1683. Los Angeles: Museum of cultural History, University of California, Los Angeles.
Straus, Lawrence Guy 1989 On early hominid use of fire. Current Anthropology 30:488-491.
Tuggle, H. David 1979 Hawaii. In The Prehistory of Polynesia. J.D. Jennings, ea., pp. 167-199. Cambridge, Massachusetts: Harvard University Press.
Van Valen, Leigh 1979 Brain size and intelligence in man. American Journal of Physical Anthropology 40:417423.
Wilson, Edward O., and William L. Brown, Jr. 1953 The subspecies concept and its taxonomic application. Systematic Zoology 2:97-111.
Woo, T.L., and G.M. Morant 1934 A biometric study of the “flatness” of the facial skeleton in man. Biometrika 26:1996-250.
Wright, Sewall 1931 Evolution in Mendelian populations. Genetics 6:97-159.
———. 1943 Isolation by distance. Genetics 28:114-138.
———. 1951 Fisher and Ford on “the Sewall Wright Effect.” American Scientist 39:452-458.
- Discuss this Article in The Multiracial Activist Forum
- Return to The Abolitionist Examiner – Home Page
Copyright © 2000 C. Loring Brace. All rights reserved.
One comment